Atsidotroofsete järvede miksotrofeerumine
Düstroofsetes ja atsidotroofsetes järvedes on biogeenid seotud enamasti humiinainetega ning pole üldjuhul fütoplanktonile ja suurtaimedele kättesaadavad. Inimtegevuse mõjul kasvava biogeenide sissevoolu tingimustes ületatakse ilmselt humiinainete biogeene siduv võime ning biogeenide sisalduse suurenemisega algab ka järve miksotrofeerumine (Joonis 23).
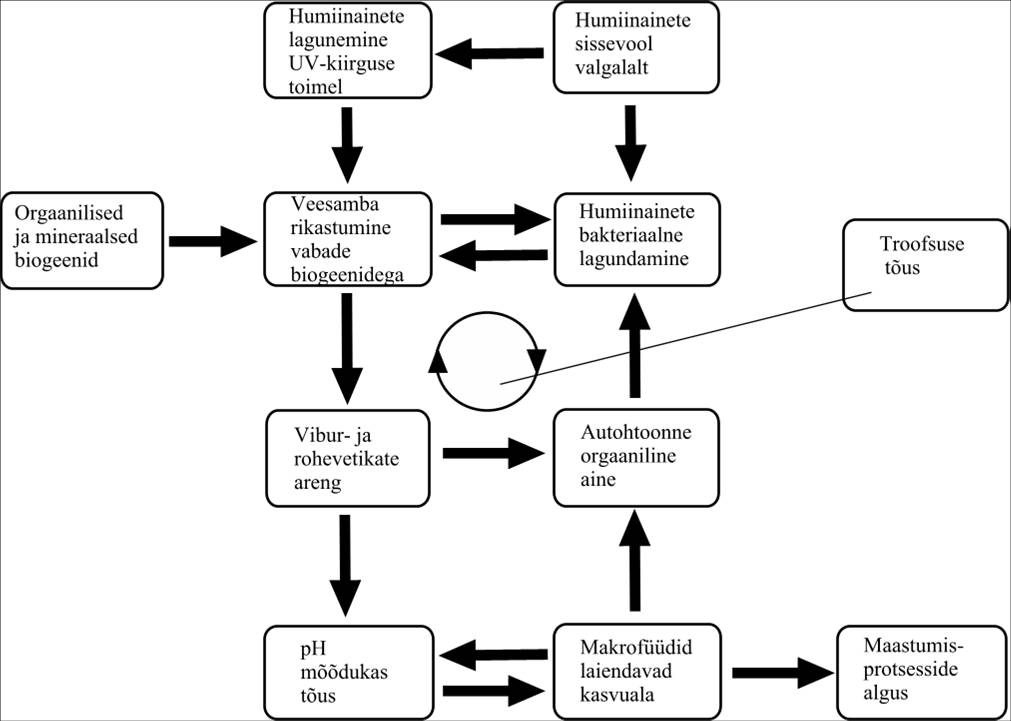
Joonis 23. Atsidotroofsete järvede miksotrofeerumine.
Lisaks otsesele kasutamisele planktoni poolt kiirendab mineraalsete biogeenide ja kergesti lagundatava orgaanilise aine lisandumine oluliselt humiinainete lagundamist, mis on düstroofsete järvede toiduahelate alglüliks. Ortofosfaatide kompleksühend raua ja humiinainetega on taaslagunev UV-kiirguse toimel, kusjuures ka humiinained ise lagunevad UV-kiirguse toimel kergemini lagundatavateks osadeks. Viimati mainitud protsess ongi üheks fütoplanktoni arengut kontrollivaks faktoriks tüüpilistes düstroofses veekogus, moodustades omalaadse biogeenide varu fütoplanktonile, olles ka ainukeseks biogeenide allikaks vähetoitelistes tumedaveelistes järvedes. Selle protsessi efektiivsust vähendab asjaolu, et humiin- ja fulvohapped absorbeerivad UV- kiirguse (lainepikkusega 200-365 nm) vaid ülemises kuni viiesentimeetrises veekihis ja vee tumedast värvusest tingitud terav ja väga püsiv kihistumine ei võimalda vegetatsiooniperioodil ulatuslikemais veemassides oleva humiinainete mõjutamist. Samas tuleb aga mainida, et suurem osa humiinainetest lagundatakse bakteriaalselt just anoksilises veekihis, allpool hüppekihti ning nii pole laguproduktid enamikule vähesest fütoplanktonist, mis paikneb soodsamate valgustingimustega veekihis, kättesaadavad. Humiinainete kolloidsete omaduste tõttu kompleksid biogeenide ja kergesti laguneva orgaanilise ainega ei setti, olles kättesaadavad kogu hüpolimnionis. Seega kontrollib fütoplankton eufootlilises kihis ning bakterid alumises veesamba osas toimuvaid protsesse. Mõnede flagellaatide nagu tativetikas (Gonyostomum semen), arengus on ilmselt olulisel kohal just eespool mainitud terav kihistus ning pinnakihtide suhteline biogeenide vaegus. Tema ohtruse kiire kasv tingitud aktiivsest migratsioonist valgustatud pinnavee ja anorgaanilise ning orgaanilise aine rikka hüpolimnioni vahel. Fakultatiivne heterotroofia ja/või vähene valgustatus võivad olla vajalikud rakkude jagunemise indutseerimiseks. Võimalik, et see liik on võimeline pagema zooplanktoni eest just sellesse veekihti ja suudab nii oma arvukust säilitada. Taolised soodsad tingimused on põhjustanud selle vetikaliigi ohtruse kasvu ja leviku laienemist Eesti järvedes.
Massiline vetikate areng loob pH mõningase tõusu tõttu soodsamad tingimused suurtaimede arenguks madalaveelistes järvedes ja lahtedes, kuid võrreldes planktoniga on ta suhteliselt inertne ja reageerib tavaliselt keskkonnamuutustele mõningase hilinemisega. Kuna makrofüütidele olulised hüdrokeemilised parameetrid nagu läbipaistvus, pH ja vee värvus muutuvad suhteliselt vähe, siis vaatamata troofsustaseme tõusule jääb suurtaimestik ikkagi suhteliselt vaeseks. Taimede suktsessioon madalates segatoitelistele veekogudele sarnastes pruuniveelistes järvedes algab ujulehtedega taimede (ujuv penikeel, vesikupp jt.) ja veesiseste taimede (tömbilehine penikeel, vesihernes) koosluste arenemisega. Hüdrofüütide risoomid muudavad turbased põhjasetted püsivamateks ja pakuvad akumulatsioonivõimalusi vees hulpivale taimsele materjalile. Selliseid paiku võivad madala vee aastatel asustada konnaosi ja laialeheline hundinui, millega algab helofüütide invasioon veekogusse. Miksotrofeerumise tulemusena suureneb düstroofsetes järvedes planktoni, suurtaimestiku ja põhjaloomastiku liigiline mitmekesisus, kuid siiski pole selge, kas kergesti laguneva orgaanilise aine ja mineraalsete biogeenide hulga vähenemisel liigub järv tagasi düstroofsesse tüüpi või jätkub autotroofse süsteemi töö juba käivitunult ning järv vananeb võrrelduna esialgse tüübiga kiiresti.
Massiline vetikate areng loob pH mõningase tõusu tõttu soodsamad tingimused suurtaimede arenguks madalaveelistes järvedes ja lahtedes, kuid võrreldes planktoniga on ta suhteliselt inertne ja reageerib tavaliselt keskkonnamuutustele mõningase hilinemisega. Kuna makrofüütidele olulised hüdrokeemilised parameetrid nagu läbipaistvus, pH ja vee värvus muutuvad suhteliselt vähe, siis vaatamata troofsustaseme tõusule jääb suurtaimestik ikkagi suhteliselt vaeseks. Taimede suktsessioon madalates segatoitelistele veekogudele sarnastes pruuniveelistes järvedes algab ujulehtedega taimede (ujuv penikeel, vesikupp jt.) ja veesiseste taimede (tömbilehine penikeel, vesihernes) koosluste arenemisega. Hüdrofüütide risoomid muudavad turbased põhjasetted püsivamateks ja pakuvad akumulatsioonivõimalusi vees hulpivale taimsele materjalile. Selliseid paiku võivad madala vee aastatel asustada konnaosi ja laialeheline hundinui, millega algab helofüütide invasioon veekogusse. Miksotrofeerumise tulemusena suureneb düstroofsetes järvedes planktoni, suurtaimestiku ja põhjaloomastiku liigiline mitmekesisus, kuid siiski pole selge, kas kergesti laguneva orgaanilise aine ja mineraalsete biogeenide hulga vähenemisel liigub järv tagasi düstroofsesse tüüpi või jätkub autotroofse süsteemi töö juba käivitunult ning järv vananeb võrrelduna esialgse tüübiga kiiresti.
